旧石器時代の特徴
🌟 基本的には打製石器(石を打ち欠いて作る)を使用
後期には一部で磨製石器も登場(特に東南アジアなど)
→ 磨製石器(石の表面を研磨して滑らかに仕上げた)は、新石器時代(約1万年前から)に世界各地で広く使用
🌟 狩猟・採集を中心に移動生活(大型動物を追う)
後期には一部で「半定住」が始まり、洞窟や岩陰を利用
世界規模での旧石器時代(約300万 ~ 1万年前)
📌約330万年前→最古の石器(ケニア・ロメクウィ遺跡)
ホモ属登場以前のヒト科(💡猿人:アウストラロピテクスなど)が使用した可能性
💡「猿人」「原人」「旧人」「新人」という分類は現在の人類学では使われなくなりつつある
現代の分類では「ホモ属(ホモ・ハビリス以降)」が人類(ヒト属)とされる
主なヒト属
世界規模での旧石器時代には複数のヒト属が同時期に存在していたがホモ・サピエンス以外は絶滅!!
現在生存しているのはホモ・サピエンス(私たち人類)だけ
| ホモ・ハビリス(約250万~180万年前) | ・最初の「ホモ属」! ・簡単な石器を使用(オルドワン石器文化) ・アフリカにのみ生息 ・ホモ・エレクトスへ進化 |
| ホモ・エレクトス(約190万~11万年前) | ・アジア・ヨーロッパへ拡散! ・ジャワ原人・北京原人など亜種を含むグループ |
| ネアンデルタール人(約40万~3万年前) | ・ ヨーロッパ・西アジアに分布し ・寒冷地に適応 ・ 高度な道具を使い、埋葬や芸術(洞窟壁画)の痕跡も! ・現代の非アフリカ系人類に2~3%のDNAが残る! |
| デニソワ人(約40万~5万年前?) | ・ シベリア・中央アジア・東南アジアの一部に分布 ・ ホモ・サピエンスやネアンデルタール人と交雑! ・ 現代のアジア・オセアニアの一部の集団にDNAが残る(最大5%) |
| ホモ・フローレシエンシス(約10万~5万年前) | ・インドネシア・フローレス島にいた小型の人類(身長約1m) ・ 脳のサイズは小さいが、石器を使用していた |
| ホモ・サピエンス(約30万年前~) | ・約30万年前にアフリカで誕生 |
📌 約7万4000年前:人類が「ほぼ絶滅しかけた」と推測されている
→ 数千〜1万人に激減
→ インドネシア・スマトラ島にある「トバ火山」が超大噴火🌋💥
→ これが地球規模の環境激変を引き起こしたと言われています
→ 現代人の遺伝子のバリエーションが少ない
→ つまり「少人数からの再スタートだった」ことを示してる
📌 約6〜7万年前:💡出アフリカ
→ ホモ・サピエンスが、アフリカを出て世界各地へ拡散
→ ネアンデルタール人やデニソワ人と交雑しながら各地に定住
出アフリカのルート
実際の出アフリカ後の拡散過程は、単純ではない
枝分かれした集団が逆流や合流を繰り返し、地域によっては隔離と交雑を経て複雑に変化
ここで示す各ルートも互いに完全に独立していたわけではない
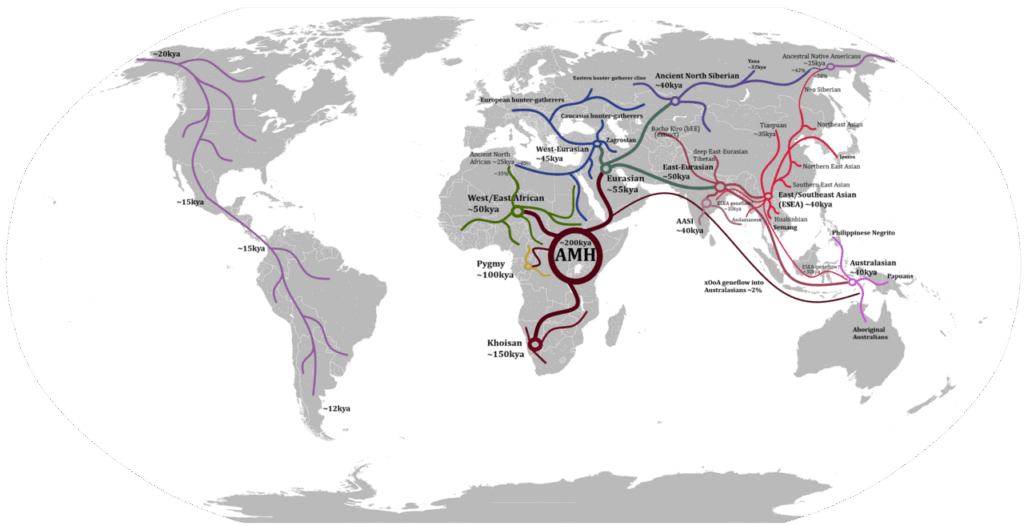
画像出典: ABCymta – 投稿者自身による著作物, map from File:Blank Map World Secondary Political Divisions.svg (2021年12月13日 11:20版), CC BY-SA 4.0, 元ページはこちら
{kind=link}
北ルートと南ルートが完全に分かれる直前、または分岐直後のごく早い段階で、ネアンデルタール人との交雑が起きている
*非アフリカ系の全人類(ヨーロッパ人、東アジア人、南アジア人、アメリカ先住民など) が、共通して約2〜3%のネアンデルタールDNAを持っている
✅南ルート:アフリカ東部 → アラビア半島沿岸 → インド → 東南アジア → オーストラリア
北ルートよりも早期(東南アジアやオーストラリアには約6万年以上前に到達)
デニソワ人と交雑(メラネシア系には約5%の遺伝子)
- アフリカ~アラビア半島(紅海横断)→海峡が狭い箇所もあったが完全に徒歩のみとは考えにくい
- インド~東南アジア本土(沿岸回廊)→氷期の海面低下により、陸続きの部分が多く、ほぼ徒歩移動が可能
- 東南アジア~オーストラリア→ 約5万年以上前にオーストラリアに人類が到達していることから、海上移動手段を持っていたと考えられる
✅ 北ルート
何度も分岐・合流・交雑を繰り返しながら、かなり複雑に広がった
| ヨーロッパへ行った人々 |
| ▲ヨーロッパへ向かった人々の一部は、さらに中央アジアやシベリア方面へ広がったものの、その後に東方から到来した別集団とも混血・再接触を繰り返したと考えられます ▲ヨーロッパ大陸への到達は約4万5千年前頃と推定され、当時まだ一部に残存していたネアンデルタール人との間で、小規模な交雑が続いた可能性も指摘されています ▲さらに、最終氷期やその後の完新世には、複数回にわたる移動・混血(たとえば新石器時代の農耕民や青銅器時代のステップ地帯からの集団流入など)も重なり、現在のヨーロッパ人集団は非常に複雑な形成過程を経てきたと考えられます |
| ヒマラヤ北側ルート(中央アジア~東アジア方面) |
| ▲出アフリカ後、ユーラシア大陸内部へと拡散した人々の一部は、中東やコーカサス・中央アジア方面を経由しつつ、ヒマラヤ山脈の北側を回り込んで東アジアに至ったとされます ▲この過程では、ステップ地帯やタリム盆地など、シルクロード沿いの要衝を通過してゆく中で、他の集団との接触・混血が繰り返され、地域ごとに多彩な遺伝的特徴が形成されました ▲さらに後代になると、シベリアから南下してくる集団や、インド亜大陸方面から北上してくる集団とも再び混ざり合うなど、単一ルートに留まらない複雑な移動の歴史が東アジア全体の遺伝的多様性をもたらしたと考えられます |
| ヒマラヤ南側ルート(南アジア方面) *完全な南ルートとは違い、北ルートから南に分岐した別ルート |
| ▲出アフリカ後にユーラシア大陸へ向かった一部の集団は、インド亜大陸の北部を回りつつ、ヒマラヤ山脈の南側から各地へと広がったと考えられています ▲このルートでは、パンジャブ平原やガンジス川流域を中心に拡散が進んだほか、時期によってはデカン高原やインド南端まで到達し、地方ごとに多彩な言語・文化・遺伝的特徴が形成されました ▲加えて、後の時代になると、イラン高原方面からの移住やアフガニスタン~インド北西部を通じた往来が繰り返され、南アジアの遺伝的構成は単一ルートの結果というより、複数の波が重なり合った複雑な経緯を経ていると考えられます |
✅アメリカ大陸へ(約2万~1.5万年前):シベリア経由でベーリング地峡(当時は陸地)を渡り、北米→南米へ移動
文化的要素
| 埋葬文化 |
| ・シャニダール遺跡(イラク) → ネアンデルタール人が埋葬し、花を添えた形跡? ・スンギール遺跡(ロシア) → ホモ・サピエンスが埋葬し、装飾品や副葬品 |
| ヴィーナス像 (📌約3.5〜2万年前) |
| ・ヴィレンドルフ(オーストリア) ・レスピューグ(フランス) ・ホーレ・フェルス(ドイツ) |
| 洞窟壁画 |
| ・ショーヴェ洞窟(📌約3.2万年前) → 最古級の動物画 ・アルタミラ洞窟(📌約1.8万年前) → 天井画が有名(宗教的意味?) ・ラスコー洞窟(📌約1.7万年前) → 狩猟やシャーマニズムと関係? |
日本での旧石器時代(約38000~16000年前)
日本列島では「旧石器時代は存在しない」と考えられていた
1949年に岩宿遺跡(群馬県)で打製石器を発見!!
旧石器時代の存在が確認された
| 石器の使用 | 打製石器が主流だが、💡磨製石器も登場 |
| 交易と流通 | 黒曜石(長野県の和田峠・北海道の白滝で産出)が、遠方の遺跡から見つかる |
| 細石器の使用 | 「後期旧石器時代」に細石器が登場 *ヨーロッパでは「中石器時代」の特徴 |
| 世界最古級の土器 | 長崎県・福井洞窟で「旧石器時代末期」に作られた土器を発見 |
💡日本で発見された約3万年以上前の磨製石器(磨製石斧)は世界最古級
日本の旧石器時代にはすでに磨製石器が使用されていた
日本列島へのルート
ホモ・サピエンスの日本列島への到達(約3.8万~3.5万年前ごろ)
| 北方ルート | 樺太 → 北海道 |
| 朝鮮半島ルート | 朝鮮半島 → 対馬 → 九州 |
| 南方ルート | 台湾 → 沖縄諸島 → 九州 |
北方ルートは陸続きだけど、朝鮮半島ルート(対馬海峡)は完全に陸続きではない(今より狭いが約50km渡海が必要)
また南方ルートの沖縄では人骨が確認されていることから
🌟航海技術があったと考えられる
| 石器 | ・約3.8万前以降の地層から多くの石器が各地で発見 → 日本列島に定住した証拠 ・金取遺跡(岩手県)で約9万年前の石器が発見されている→もし人工品なら、旧人がいた可能性も❓❓ |
| 人骨 | 📌約32000年前→山下洞人(沖縄県) 📌約27000年前→白保竿根田原洞穴(沖縄県) 📌約20000年前→港川人(沖縄県) 📌約14000年前→浜北人(静岡県) |
現代日本人のDNAのこと
✅ミトコンドリアDNAとY染色体
| ミトコンドリアDNA | Y染色体 |
| 母 → 子(性別問わず) | 父 → 息子のみ |
| 男性は次に伝えられない | 女性にはY染色体はない |
ミトコンドリアDNA・Y染色体は、一直線で受け継がれる
→ 突然変異を調べることで系統を分類できる
→ これがハプログループ
→ 男系のルーツを知りたいときは「Y染色体ハプログループ」
→ 母系のルーツを知りたいときは「mtDNAハプログループ」
✅核DNA(常染色体)
→ 父母から半分ずつランダムに混ざって受け継がれる
→ どれくらい似てるかの相関関係
Y染色体 vs ミトコンドリア vs 常染色体
| Y染色体ハプログループ | mtDNAハプログループ | 核DNA(常染色体) |
| 男系のルーツ | 母系のルーツ | 父・母から半分ずつ |
| 男性のみ調べられる | 男女OK | 男女OK |
✅ 日本人男性に多いハプログループ
| D1b(旧称D2):約30〜40% | 特に本州の山間部やアイヌに多い 縄文系・1万年以上前から列島にいたと考えられる古い系統 |
| O1b2:約30〜40% | 日本の主流Y染色体(都市部に多い) 弥生系 → 稲作とともに朝鮮半島〜中国江南地方から来たとされる |
| O2:約10〜15 | 弥生時代後半〜古墳時代以降に流入? 中国大陸南部から |
| C1a1:1〜2%未満 | 縄文時代にいたけど、ほとんど絶滅した古いグループ |
| N系統:1%未満 | シベリアや北方アジア経由の少数派ルート |
👉 D1b(縄文系)は、日本以外ほぼ存在しない超レア系統!
チベットやアンダマン諸島に「D系統」の親戚はいるけど、D1bはほぼ日本限定!
✅ DNA(常染色体)では
🟤 縄文人系 約10〜20% 1万年以上前から日本にいた先住民(狩猟採集民)
🟡 弥生人系 約70〜80% 稲作文化とともに大陸から来た(中国〜朝鮮半島系)
🔵 その他(渡来系) 数%程度 古墳時代〜中世にかけての移民(ツングース系、漢族など)
現代日本人のゲノムを解析し、縄文人由来の遺伝的変異の地域差を調査した結果
東京大学の渡部裕介特任助教と大橋順教授らによるもので、2023年3月に科学誌 iScienceに発表
出典:渡部裕介・大橋順「Modern Japanese ancestry-derived variants reveal the formation process of the current Japanese regional gradations.」(iScience, 2023年3月)
日本経済新聞「日本人のルーツ、ゲノムで地域差がくっきり」(2023年12月22日)
| 最も少ない(0.0158~0.0164) | 愛媛・高知・香川・徳島 和歌山・奈良・三重・滋賀・福井 |
| 最も多い県(0.0184~0.0195) | 鹿児島・島根 群馬・茨城・福島・宮城・岩手・秋田・青森 |
✅ 日本人のルーツ
→ 従来は、二重構造(縄文+弥生)
→ 近年のゲノム解析から、より複雑な背景をもち、複数のルートを通じて列島に到達していた
→ 実は3層構造だった(縄文+弥生+古墳以降)ことが示唆されてる
地球環境の変化
気候の変化(温暖化)が暮らしを変え、新たな時代へ(新石器時代へ移行)
更新世(氷河時代)と完新世
| 更新世(氷河時代) | 完新世 |
| 約258万~11700年前まで | 約11700年前~現在 |
| 氷期と間氷期を繰り返す 最終氷期(約11万~11700年前)が終わると、温暖化が進み完新世へ | 最終氷期が終わり、比較的温暖で安定した気候が続く |
温暖化の影響
| 世界(メソポタミア・中国など) | 日本 |
| 新石器時代へ | 縄文時代へ(独自の文化) |
| 温暖化 → 農耕・牧畜へ | 温暖化 → 豊かな自然で狩猟採集を継続 |
| 定住化+農耕・牧畜(生産経済) | 定住化+狩猟採集(獲得経済のまま) |
約1.2万年前、氷河期が終わり気温が上昇
森林が拡大し、植物や動物の分布が変化。大型動物(マンモス・ナウマンゾウなど)は絶滅し、狩猟中心の生活が難しくなる
世界では農耕が始まり(新石器時代)、日本では豊かな自然を活かした狩猟採集+定住(縄文時代)へ
*最古級の縄文土器は温暖化以前(氷河期末期)に存在した
また、氷床が溶けて海面が上昇し、日本列島は大陸から完全に切り離された
🕓 更新日:2025年7月6日
※ 個人的な備忘録として、調べながら書いているブログで、書き足し、修正、アップデートを重ねています